02 – Taxonomia, mineralogia composicional e considerações sobre ambientes de fossilização dos ictiolitos da Formação Santana (Eo-Cretáceo da Chapada do Araripe): estudo de caso do acervo didático do Laboratório de Paleontologia da Faculdade de Geologia da Universidade Federal do Pará.
Ano 04 (2017) - Número 03 Artigos
![]() 10.31419/ISSN.2594-942X.v42017i3a2BESB
10.31419/ISSN.2594-942X.v42017i3a2BESB
Breno Eduardo da Silva Barros1; José Fernando Pina Assis2; Horácio Higuchi3
Resumo
Este trabalho trata da identificação taxonômica de representantes da paleoictiofauna da Formação Santana (Eo-Cretáceo marinho do Brasil), procedentes da região da Chapada do Araripe, Estado do Ceará e pertencentes ao acervo didático do Laboratório de Paleontologia do Departamento de Geologia da Universidade Federal do Pará (UFPA). Foram analisadas trinta e uma amostras da coleção e identificados sete gêneros (Calamopleurus Agassiz, 1841; Vinctifer Jordan,1919; Cladocyclus Agassiz, 1841; Rhacolepis Agassiz, 1841; Notelops Woodward, 1901; Dastilbe Jordan, 1910; Tharrhias Jordan & Branner, 1908. Além disso, considerando-se o excelente estado de conservação dos espécimes, foram realizadas análises mineralógicas com o emprego de microscopia eletrônica de varredura (MEV) em onze amostras selecionadas, no sentido de obter maiores detalhes sobre a estrutura composicional de vértebras e escamas, bem como foram tecidas considerações sobre paleoambientes reinantes à época dos processos de fossilização.
Introdução
O acervo fossilífero do Laboratório de Paleontologia do antigo Departamento de Geologia (atual Faculdade de Geologia) da Universidade Federal do Pará abriga expressiva coleção de concreções ictiolíticas, procedente da Formação Santana (Chapada do Araripe, Estado do Ceará). São mais de 50 peças, em sua maioria, contendo exemplares incompletos, incluindo corpos, partes caudais, centrais e craniais, absolutamente todas em excelente estado de conservação.
A despeito disso, o material nunca havia sido objeto de um trabalho específico de identificação taxonômica, sendo utilizado apenas nas atividades didáticas das disciplinas dos cursos de graduação em geologia e biologia da universidade.
Em meados de 2002 o então acadêmico Breno Eduardo da Silva Barros, concluinte do Curso de Licenciatura em Ciências Biológicas da UFPA assumiu a responsabilidade pela realização de seu Trabalho de Conclusão de Curso (TCC) enfocando a coleção mencionada, sob orientação do prof. José Fernando Pina Assis e co-orientação do Dr. Horácio Higushi, pesquisador do Museu Paraense Emílio Goeldi (MPEG/CNPq).
Este trabalho apresenta os resultados obtidos com a realização do TCC: identificação taxonômica da coleção, mineralogia composicional de vértebras e escamas, além de breves considerações sobre possíveis condições ambientais de fossilização.
Aspectos Geológicos da Chapada do Araripe
Trata-se de uma região compreendida entre os estados do Ceará, Piauí e Pernambuco (Figura 1), relativamente elevada e plana, alongada em sentido leste-oeste, medindo quase 180 km e cerca de 50km em sentido transversal (Figura1). Em sua borda oriental, a altitude ultrapassa 950m, enquanto que próximo à fronteira com o Piauí é de cerca de 700m (Brito, 1990, Silva Santos, 1991). A Chapada do Araripe é divisor de águas entre rios Jaguaribe e São Francisco.
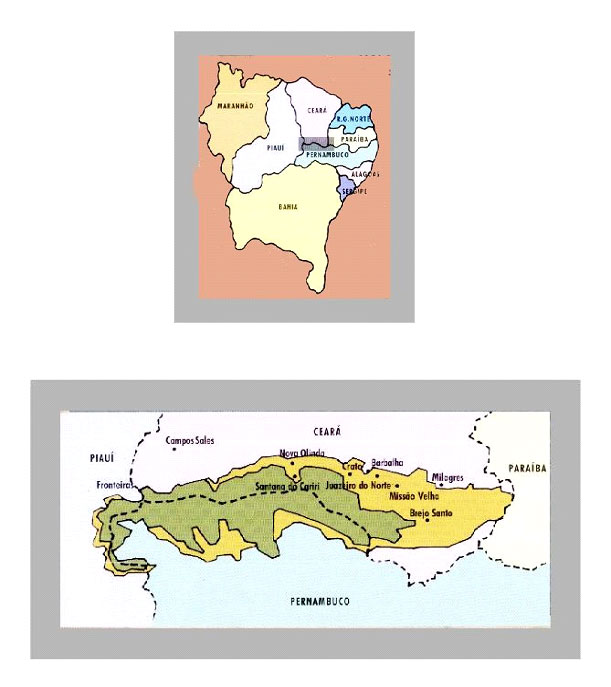
Figura 1 – Localização geográfica da Chapada do Araripe no contexto da região nordeste do Brasil. Modificado de Museu de Paleontologia/Universidade Regional do Cariri – folder.
A chapada é testemunho dos processos de modelamento da paisagem, atuantes sobre uma espessa seqüência sedimentar paleozoico-mesozóica, depositada sobre embasamento cristalino. Sua área geológica transcende os limites da própria chapada, porém é nos paredões escarpados que se pode examinar seus estratos (Silva Santos, 1991). Segundo Beurlen (1971), a seqüência sedimentar da Chapada do Araripe representa uma ocorrência isolada, não ligada a qualquer outra unidade sedimentar, repousando sobre o bloco cristalino brasileiro.
Estratigraficamente compreende cinco unidades: Formação Cariri, Formação Brejo Santo, Formação Missão Velha, Formação Exú e Formação Santana, esta última, devido ao seu surpreendentemente rico conteúdo fossilífero, no qual abundam peixes, é objeto de vários trabalhos de cunho paleontológicos e geológicos (Brito, 1990). As camadas sedimentares são praticamente horizontais, com ligeiro mergulho para oeste na encosta oriental, sobrepostas aos sedimentos detríticos paleozóicos da Formação Cariri, correlacionáveis aos arenitos silurianos da Formação Serra Grande (da Bacia do Parnaíba).
A Formação Santana
Foi criada por Small (1913), que a denominou Calcáreo de Sant’Anna. Beurlen (1962) retrabalhando a litoestratigrafia regional, subdividiu-a em três membros: 1) calcários inferiores laminados; 2) gipsita; 3) calcários superiores margosos. Em 1963 o mesmo autor criou a Formação Crato, para englobar os calcários superiores descritos no ano anterior.
Mabessone e Tinoco (1973) baseando-se em parâmetros paleoambientais, subdividiram a formação novamente em três membros: Membro Crato (basal), constituído de calcário clástico e quartzoso; Membro Ipubi (intermediário), com os níveis de Gipsita e Membro Romualdo (superior), constituído de camadas espessas de calcário. Lima (1978), com base em dados palinológicos confirmou a existência das três fases distintas de deposição, como foram propostas por Beurlen em 1971. Santos (1982), num amplo trabalho de reconhecimento sobre o ambiente deposicional da Formação Santana, identificou diversos litofacies e encontrou restos de vegetais nos calcários laminados (algas filamentosas e oogônios de carófitas) além de conchostráceos, ostracodes, insetos e peixes, que apontaram um ambiente dulciaqüícola para a unidade.
Campos e Wenz (1982), descrevendo peixes celacantídeos na Chapada do Araripe, resumiram a coluna estratigráfica, corroborando a subdivisão da Formação Santana no três membros de Mabessone e Tinoco (op. cit.). O membro Crato, constituído de calcários e folhelhos, apresenta insetos terrestres e aquáticos, bem como outros artrópodes, além de vários vertebrados, incluindo diversos peixes, um sapo e uma pena, sendo considerado como produto de ambiente lacustrino. O membro Ipubi, constituído de evaporitos, (principalmente Gipsita) e sem registros de fósseis, é o resultado de deposição em ambiente com significativo aporte de sais. O membro Romualdo, constituído de folhelhos, margas e calcários, apresenta em sua base a indicação de ambiente continental, enquanto no topo já teria sido depositado em ambiente marinho, conforme indicado pela presença de equinóides e vários grupos de peixes marinhos, onde se encontram as concreções calcárias (Maisey, 1991; Maisey in Goldblatt, 1993).
O Registro Fóssil da Formação Santana Apresenta conteúdo abundante: Entomofauna com aproximadamente duzentas espécies descritas (Brito, 1984; Martins-Neto et al, 1991, 1992; Maisey, 1991); Herpetofauna com espécies de pterossauros distribuídas em quatro famílias, várias espécies de dinossauros, duas espécies de quelônios e duas espécies de crocodilianos segundo Maisey, (1991); Ornitofauna com uma raríssima ocorrência de penas fossilizadas, cujas condições se dão apenas em águas calmas, de ambientes lacustrinos ou lagunares (Martins Neto e Kellner, 1988 in Maisey, 1991); Carcinofauna registra a ocorrência dos conchostráceos Estheria e Bairdestheria, principalmente no membro Romualdo segundo Lima (1979). Esse conjunto fóssil abrangente e extremamente determinante, para a reconstrução paleoambiental do Cretáceo mundial. Sua paleobiota impressiona pela riqueza e abundância de formas, com registros fósseis que vão, desde vegetais até vertebrados alados, passando por sua riquíssima paleoictiofauna, talvez o mais expressivo registro fóssil mundial, considerando-se seu intervalo de tempo.
A Ictiofauna
O primeiro registro descrito na região data de 1817, fruto das expedições realizadas pelos naturalistas Johann B. von Spix e Carl F. Phil von Martius, que acreditavam tratar-se o espécime de uma serpente. Hoje, a partir das ilustrações feitas pelos autores, acredita-se ser na verdade um peixe do gênero Rhacolepis Agassiz,1841. Infelizmente, o exemplar perdeu-se no decorrer dos anos.
Os peixes fósseis da Chapada do Araripe têm enorme importância para o entendimento do processo evolutivo dos peixes, na medida em que incluem representantes que marcam a transição dos Holostei para os Teleostei, num processo de ossificação das estruturas, típico destes últimos. Também marcam o gradativo desaparecimento das escamas ganóides, características dos peixes holósteos, fenômeno iniciado no Período Jurássico e conhecido como declínio dos holósteos. A paleoictiofáuna do Araripe, em especial os Actinopterygii, representa os últimos estádios evolutivos dos Osteichthyes (Carroll, 1988; Silva Santos, 1991). Segundo Maisey (1991), já foram descritos vinte e um gêneros, distribuídos em quinze famílias, além Araripelepidotes, ainda sem posição sistemática definitiva, e um clupeomorfídeo, não identificado.
A Coleção do Laboratório de Paleontologia é composta por 86 exemplares de concreções ictiolíticas calcárias, a maioria fragmentos das regiões toráxicas, caudais e craniais, todas em excelente estado de conservação. Para o trabalho taxonômico foram selecionadas 31 peças (Tabela 1), consideradas representativas da coleção. Em virtude do objetivo a ser atingido na análise de cada amostra, os procedimentos laboratoriais seguiram como abaixo:
Tratamento Mecânico – lavagem em água corrente e escovação com cerdas finas para retirada de restos de calcário que recobriam as regiões objeto de análise. Separação em grupos, considerando a morfologia das concreções.
Tratamento Químico – lavagem das peças em meio ácido para melhorar a visualização de estruturas diagnósticas. As amostras V-515 (Cladocyclus), 455-V (Vinctifer) e V-506 (Tharrhias) foram submetidas ao ataque com ácido clorídrico (HCl) diluído a 10%, para clareamento químico e identificação de estruturas ósseas da matriz em calcário. O ataque foi feito sobre cada amostra durante duas semanas com pincel de cerdas plásticas, intercalando-se lavagem em água corrente após cada sessão de pincelagem.
Análise em Lupa Binocular – As amostras V-501 (Dastilbe); 454-V (Vinctifer); V-513 (Tharrhias); V-516, 527-V (Rhacolepis); V-515, 559-V (Cladocyclus) e S-230 (Notelops), foram analisadas em lupa estereoscópica Zeiss, com aumento de 10X, para identificação mais detalhada das vértebras, estruturas craniais, e escamas.
Documentação Fotográfica – foram obtidas com câmera fotográfica Nikkon FX-3, lente macro-zoom 70-160mm e câmara digital Sony Mavica, MVC-FD73, sobre mesa estativa, com luz dirigida em rampa a 25o de inclinação, em virtude dos baixos relevo e contraste de cor das peças.
Documentação em Desenho – de autoria de Breno Eduardo Barros, foi feita para complementar a identificação de peças osteológicas pouco perceptíveis nas fotografias. Os desenhos foram elaborados com lápis (grafite 3B) e posterior cobertura em nanquim, sem o auxílio de câmara clara. As peças com estruturas mais visíveis ou melhor preservadas, compõem as estampas do trabalho.
Tabela 1 – Relação das amostras estudadas e suas características morfológicas.
| Gênero | N.º de registro | Características da amostra |
|---|---|---|
| Calamopleurus | 437-V | Impressão externa do tronco do peixe, fragmentado |
| CA | Impressão externa do tronco do peixe, fragmentado | |
| Vinctifer | V – 528 | Impressão externa do tronco e crânio |
| 469 – V | Impressão externa do tronco | |
| 442 – V | Fragmento do indivíduo, tronco e crânio | |
| 444-V | Impressão externa do tronco | |
| 455 – V | Fragmento do indivíduo, tronco | |
| 456 – V | Fragmento do indivíduo, frag. de tronco e crânio fragmentado | |
| V – 517a | Frag. do indivíduo, frag. de tronco e crânio fragmentado | |
| V – 517b | Impressão externa, frag. de tronco e crânio fragmentado | |
| V – 517c | Fragmento do indivíduo, tronco | |
| V – 517d | Impressão externa do tronco | |
| 454 – v | Fragmento do indivíduo, crânio | |
| V – 560 | Fragmento do indivíduo, tronco e crânio | |
| VA | Impressão externa do tronco e crânio | |
| VB | Fragmento do indivíduo, tronco | |
| VC | Impressão externa do tronco | |
| VD | Fragmento do indivíduo, tronco | |
| VE | Impressão externa do tronco | |
| Cladocyclus | 559 –V | Frag. do indivíduo, frag. de tronco e crânio |
| V–515 | Frag. do indivíduo e molde externo, frag. de tronco e crânio | |
| Rhacolepis | 527–V | Fragmento do indivíduo, fragmento de tronco e crânio |
| V – 516 | Fragmento do indivíduo, fragmento de tronco e crânio | |
| Notelops | S – 230 | Fragmento do indivíduo e impressão externa do crânio |
| Dastilbe | V–501 | Fragmento do indivíduo, indivíduo completo |
| Tharrias | V–503 | Molde externo, indivíduo completo |
| V–513 | Fragmento do indivíduo, indivíduo completo | |
| 530–V | Molde externo, fragmento de tronco e crânio | |
| 524–V | Frag. do indivíduo + molde externo, tronco e crânio | |
| 505–V | Fragmento do indivíduo, indivíduo completo | |
| TA | Molde externo, fragmento de tronco e crânio |
Classificação Taxonômica, segundo Wenz & Brito (1990) e Maisey (1991).
Divisão HALECOSTOMI
Subdivisão HALECOMORPHI
Família AMIIDAE Bonaparte, 1837
Subfamília AMIINAE Patterson, 1973
Calamopleurus Agassiz, 1841
(Figuras 2 e 3)

Figura 2 – Impressão da parte interna de um tronco de Calamopleurus, em concreção do tipo 2 (Maisey,1991); registrada sob número 437-V. destaque para 8 impressões de vértebras e 4 corpos vertebrais preservados.
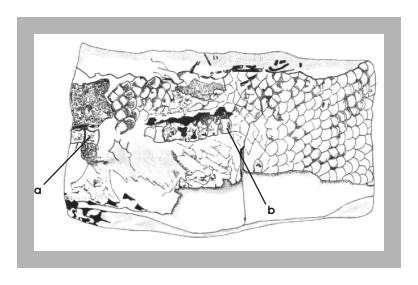
Figura 3 – Desenho esquemático figura anterior, destacando-se impressões das vértebras (a) e corpos vertebrais (b).
Subdivisão TELEOSTEI
Família ASPIDORHYNCHIDAE Nicholson e Lydekker, 1889
Vinctifer Jordan, 1919
(Figuras 4, 5, 6 e 7)

Figura 4 – Crânio de Vinctifer em concreção do tipo 1 (Maisey,1991), medindo 6,60 cm de comprimento, registrado sob número 454-VV.
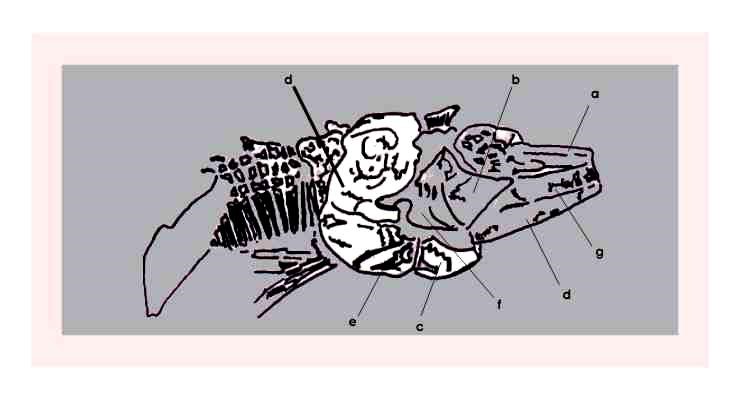
Figura 5 – Desenho esquemático da figura anterior: a) rostro-dermoetmóide; b) sub-orbitais; c) pré-opérculo; d) opérculo; e) sub-opérculo; f) hiomandibular; g) maxilar; e h) dentário.

Figura 6 – Crânio de Vinctifer, em concreção do tipo 3 (Maisey, 1991). medindo 17,2 cm, registrado sob número 560-V e exposto no Museu de Geociências da Universidade Federal do Pará. Exemplar cedido pelo Prof. Dr. Marcondes Lima da Costa.
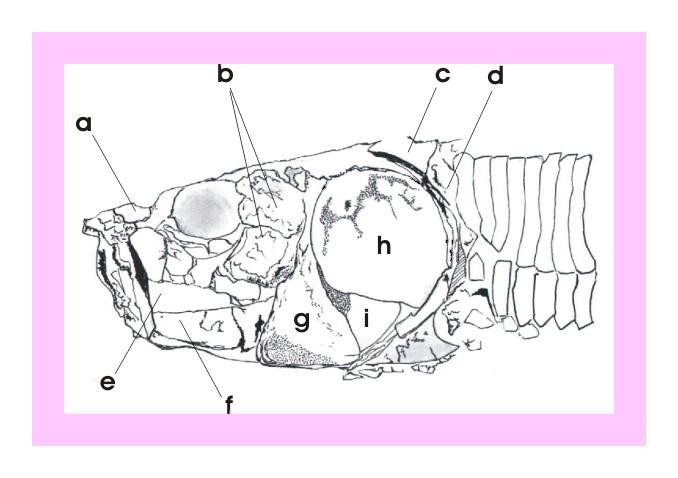
Figura 7 – Desenho esquemático da figura anterior: a) fragmento do osso rostro-dermoetmóide; b) suborbitais; c) fragmento do osso supratemporal; d) cleitro; e) fragmento do maxilar; f) fragmento do osso dentário; g) pré-opercular; h) opérculo; e i) sub-opercular.
Ordem ICHTHYODECTIFORMES (Bardack & Sprinkle, 1969)
Sub-ordem ICHTHYODECTOIDEI (Romer, 1966)
Família CLADOCYCLIDAE
Cladocyclus Agassiz, 1841
(Figuras 8 e 9)

Figura 8 – Exemplar de Cladocyclus, em concreção do tipo 4 (Maisey, 1991). medindo 5,14 cm de comprimento, registrado sob número 515-V.
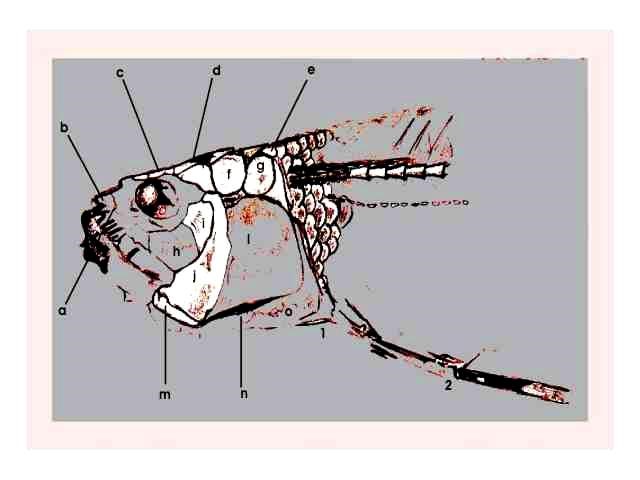
Figura 9 – Desenho esquemático da figura anterior; a) osso dentário, b) osso pré-maxilar, c) osso frontal, d) Parietal, e) crista supra-occipital, f) supra-temporal, g) pós-temporal, h) 3º inter-orbital, i) 4º inter-orbital, j) osso pré-opercular, l) opérculo, m) osso inter-opercular, n) osso sub-opercular, o) osso do cleitro.
Família PACHYRHIZODONTIDAE
Rhacolepis Agassiz 1841
(Figuras 10 e 11)
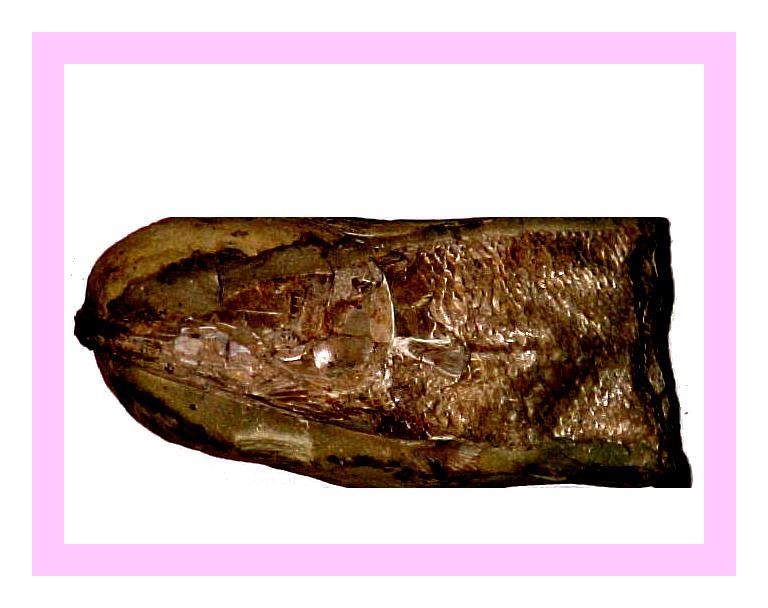
Figura 10 – Fragmento de tronco e parte do crânio de Rhacolepis, em concreção do tipo 4 (Maisey, 1991). medindo 11 cm, registrada sob número 527-V.
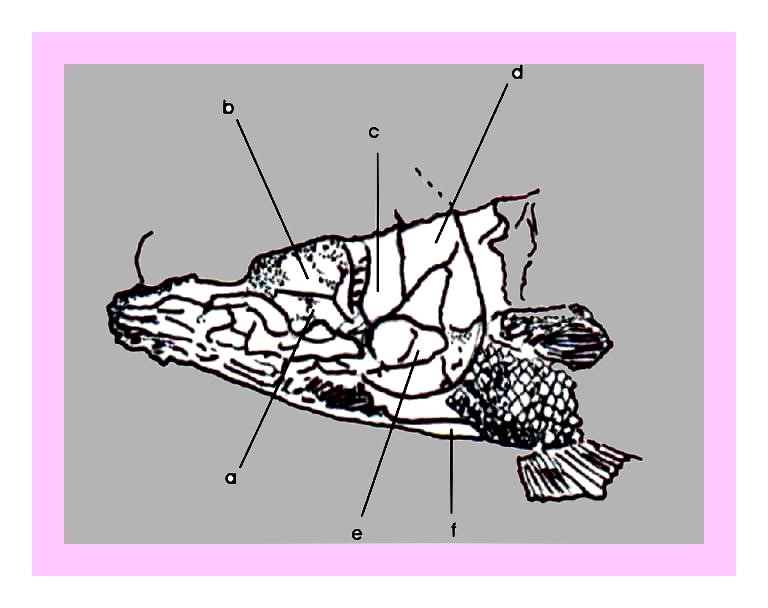
Figura 11 – Desenho esquemático da figura anterior: a) 2º inter-orbital, b) 3º inter-orbital, c) osso pré-opercular, d) opérculo, e) osso sub-opercular esquerdo, f) osso sub-opercular direito.
Família NOTELOPIDAE Forey 1977
Notelops Woodward 1901
(Figuras 12 e 13)

Figura 12 – Fragmento de crânio de Notelops, em concreção do tipo 4, (Maisey, 1991), medindo 11,5 cm, registrado sob número S-230.
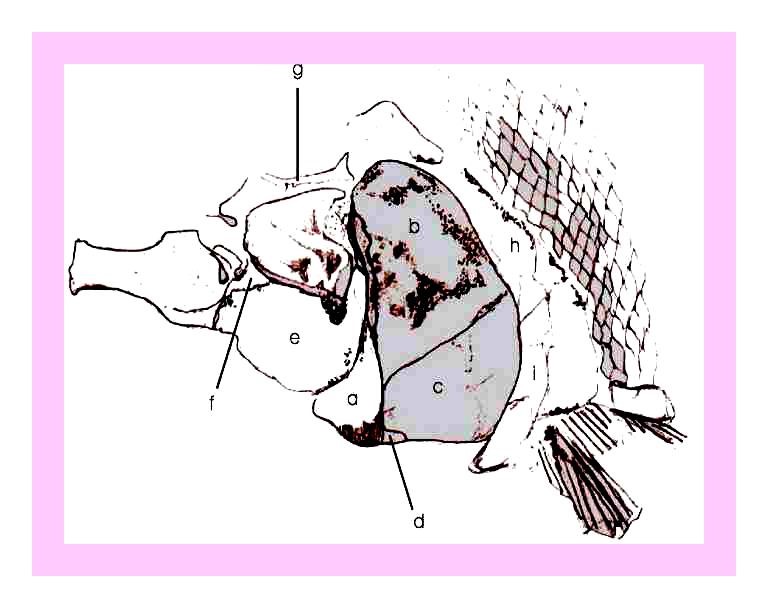
Figura 13 – Desenho esquemático da figura anterior: a) osso pré-opercular; b) opérculo; c) sub-opérculo; d) osso inter-opercular; e) 2º inter-orbital; f) 1º inter-orbital; e g) Fragmento do osso frontal.
Ordem GONORHYNCHIFORMES incertae sedis
Família CHANIDAE
Dastilbe Jordan, 1910
(Figuras 14 e 15)

Figura 14 – Molde externo do único exemplar de Dastilbe pertencente à coleção, medindo 3,2 cm e registrado sob número 501-V.
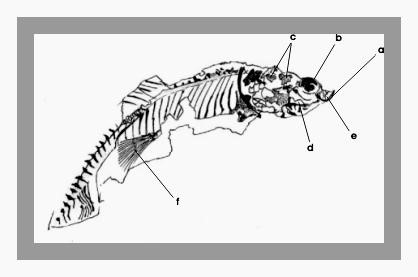
Figura 15 – Desenho esquemático da figura anterior: a) fragmento do osso dentário, b) fragmento do osso frontal, c) dois fragmentos do osso hiomandibular, d) fragmento do osso metapterigóide, e) fragmento do osso maxilar, e f) nadadeira ventral.
Família CHANIDAE
Tharrhias Jordan & Branner, 1908
(Figuras 16 e 17)

Figura 16 – Exemplar integral de Tharrhias, em concreção do tipo 3 (Maisey, 1991). medindo 13,4 cm, registrado sob número V-513.
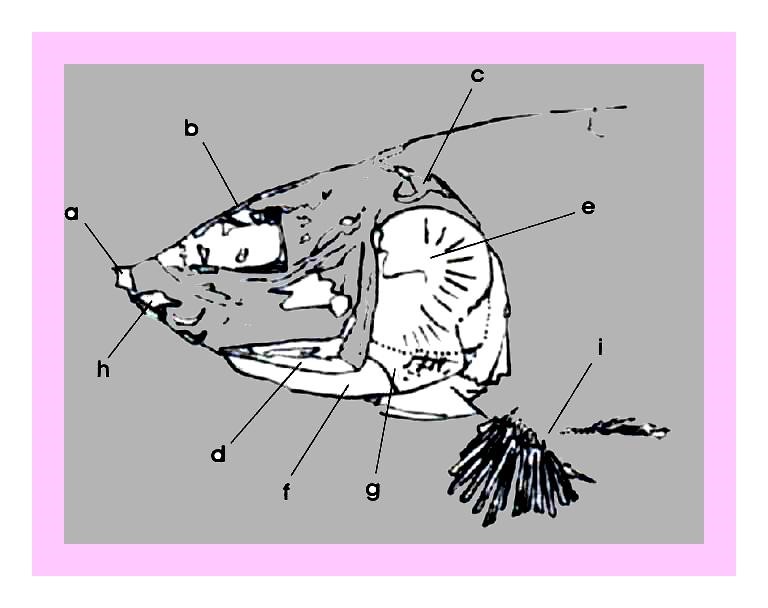
Figura 17 – Desenho esquemático da figura anterior mostrando crânio e pós-crânio imediato: a) fragmento do osso pré-maxilar, b) fragmento do osso frontal, c) fragmento do osso pós-temporal, d) osso pré-opercular, e) opérculo, f) osso inter-opercular, g) Osso sub-opercular, h) fragmento do osso dentário, i) cintura escapular.
Mineralogia
O emprego do microscópio eletrônico de varredura (MEV) como auxílio no trabalho de identificação de estruturas e composição química de peças fossilizadas tem sido estimulado nos últimos anos. Como o objetivo de estabelecer possíveis parâmetros paleoambentais, algumas amostras contendo vértebras e escamas do material da coleção foram selecionadas e submetidas a análises no Laboratório de Microscopia Eletrônica de Varredura (MEV) do Museu Paraense Emílio Goeldi, utilizando de um equipamento modelo LEO 1450-VP, sob orientação do Dr. Hilton Tulio Curtis.
Um dos aspectos que permite estabelecer critérios para identificação de processos diagenéticos é o da mineralogia restrita, entendida aí como a composição mineralógica das partes esqueletais dos restos preservados. Esqueletos de organismos vertebrados são em geral compostos por uma mineralogia muito específica, via de regra, limitada aos fosfatos, a exemplo da apatita. Procedimentos similar foi realizado por Costa (2002) em fragmentos de ossos fósseis das megafauna no Acre, em que a fluorapatita é o mineral principal, com modificações diagenéticas, mas sendo o mineral reliquiar dos tecidos ósseos.
Bonfim Junior (2001) apresentou um estudo muito interessante a respeito de constituição mineralógica de estruturas esqueletais em vertebrados, a partir do exame de dentes e ossos. Segundo ele, o tecido ósseo dos amniotas é constituído basicamente por hidroxi-apatita [Ca5(PO4)3OH] sendo muito comum a presença ainda de fluorapatita, cloroapatita e carboapatita.
Durante o processo de fossilização a hidroxi-apatita original do tecido ósseo pode se transformar em fluorapatita [Ca5(PO4)3F], fator que favorece, não somente a preservação dos ossos (as chamadas partes duras) como também a fosfatização de tecidos (as chamadas partes moles). Ainda segundo Bonfim Junior (2001), a mineralização nos ossos e dentes é gerada a partir da síntese feita por células do mesênquima, que depositam os minerais gerados em uma matriz fibrosa de colágeno. Isto cria uma estrutura cristalina especial, na qual os cristais, influenciados pelos escleroblastos, formarão lamelas mineralizadas de fibras colágenas, arranjadas concentricamente ao redor dos canais, formando o sistema haversiano dos ossos e dentes.
Nosso trabalho analisa a composição mineralógica de 11 peças destacadas dos exemplares da coleção, entre fragmentos de placas de escamas e fragmentos de corpos vertebrais, a partir das análises químicas obtidas por EDS/ MEV (Espectrometria de raios X de Energia Dispersiva acoplada ao microscópio eletrônica de varredura), com o propósito de experimentar algumas idéias de interpretação para os processos possivelmente ocorridos durante e/ou logo após o soterramento da biota do Araripe.
Uma dessas idéias advém da identificação dos principais minerais que se formaram nestas concreções e que processos de mineralização podem ter ocorrido ao longo da formação dos fósseis. Cada imagem gerada pelo MEV possibilitou a escolha de vários pontos representativos. Com o uso do detector de elétrons retroespalhados, (backscattered eletrons) foi possível obter imagens de boa resolução além de análises químicas qualitativas, que combinados permitiram inferir a composição mineralógica. De acordo com a metodologia empregada, quanto mais escura for a tonalidade do contraste gerado na imagem, mais leve é o peso atômico do elemento que compõe aquele ponto da amostra, e quanto mais clara a tonalidade, mais pesado é o peso atômico do elemento. Para essas análises foram selecionadas 12 peças da coleção, conforme discriminadas a seguir:
- VA (1SR) – escamas de Vinctifer (sem registro na coleção);
- SR – fragmento ósseo de Tharrhias (sem registro na coleção);
- 448–V – escamas de Rhacolepis;
- 449-V – escamas de Rhacolepis;
- V-512 – escama de Vinctifer;
- V-517 – placa de escama de Victifer;
- 559-V – fragmento de vértebra de Cladocyclus;
- S-230 – fragmento de vértebra de Notelops;
- 437-V – fragmento de vértebra de Calamopleurus;
- CA (NS)– fragmento de vértebra de Calamopleurus.
- VD (“d”) – escamas de Vinctifer; um fragmento de vértebra;
- V-516 – vértebras de Rhacolepis; um fragmento com escamas;
Foram analisados 31 pontos, considerados representativos das 12 peças. Em 25 deles, foram identificados os seguintes elementos com seus valores médios entre parênteses: cálcio (28,9%), fósforo (14,1%) e oxigênio (46,1%), totalizando 89,1%; os 10,1 % restantes estão representados alumínio, magnésio, silício, ferro, manganês, sódio, cloro e bário. Essa constituição química sugere o domínio de apatita, ou seja, que a mesma representa claramente a manutenção das estruturas originais produzidas pelos organismos, sendo a apatita o mineral comum na estrutura óssea.
A presença de barita, identificada nas amostras 516 V/6, 517 V/1 e D/2 1, (Figuras 18 e 19) sugere condições de substituição dos compostos fosfatados por compostos sulfatados durante a fase diagenética, o que leva a um questionamento sobre as condições de oxidação ou anoxidade do meio, durante a fase estratinômica.

Figura 18 – Micrografia eletrônica de corpo de vértebra de Rhacolepis, (amostra 516-V), obtida com detector de elétrons retro-espalhados. O ponto marcado como 516 V/2 5 é um dos pontos onde foi realizada análise com o EDS.

Figura 19 – Detalhe da micrografia anterior, mostrando cristal de barita (BaSO4).
A esse respeito, Maisey (1991) menciona Schaffer (1972), Seilacher et al (1985), como indicadores de que os peixes da Formação Santana fossilizaram em meio hipersalino, baixa concentração de oxigênio e elevada aridez, fator que teria permitido a rápida evaporação da água, aumentando assim a concentração dos sais. Nestas condições, os sulfatos encontram ambiente ideal para precipitar. Assim, não é de estranhar a presença de barita nas estruturas ósseas. Sufossal de bário, esse mineral demanda condições oxidantes para sua formação. As análises de EDS sugerem a presença de barita (Figura 20), o que permitiu concluir que durante a fase de soterramento, o ambiente químico indicava condições ainda oxigenantes, porém em nível de troca iônica elevado.
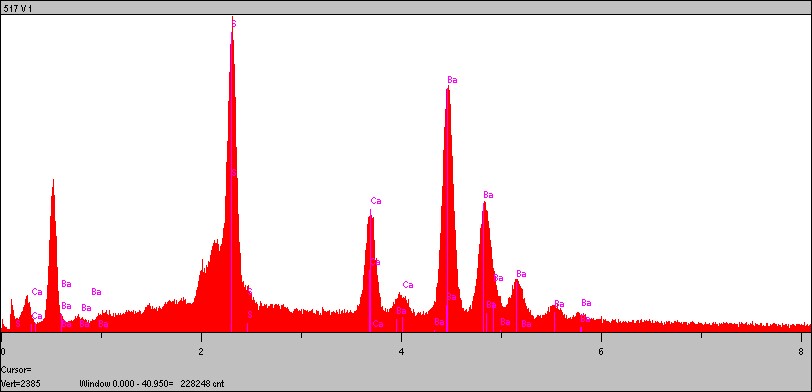
Figura 20 – Espectro de análises químicas EDS/MEV mostrando o domínio de S, Ba e Ca encontrados no fragmento de escama de Vinctifer no ponto 517 V/1. Essa constituição química sugere a presença de barita como mineral principal.
Conclusões
Nosso trabalho permitiu a identificação taxonômica de sete gêneros: Vinctifer Jordan, 1920, (17 peças), Calamopleurus Agassiz, 1841 (2 peças), Cladocyclus Agassiz, 1841 (2 peças); Rhacolepis Agassiz, 1841 (2 peças), Tharrhias Jordan & Branner, 1908 (6 peças), Notelops Woodward, 1901 e Dastilbe Santos, 1948 (ambos com 1 peça), totalizando 31 amostras selecionadas no acervo do Laboratório de Paleontologia do Departamento de Geologia da UFPA.
O trabalho destaca a importância da ictiofauna da Chapada do Araripe para a compreensão de aspectos evolutivos dos peixes ósseos. A presença de representantes Holostei (Calamopleurus e Vinctifer) e representantes Teleostei (Cladocyclus e Notelops) nos mesmos habitats, permite pensar que disputavam os mesmos nichos ecológicos. Além disso, também foi observada a passagem de alguns caracteres para outros, como escamas ganóides (dos Holostei), gradando para escamas ciclóides (dos Teleostei primitivos). Por tudo isso e como apontado por Carroll (1988) e Silva Santos (1991), a paleoictiofauna da Chapada do Araripe, notadamente os Actinopterygii, representa os últimos estádios evolutivos dos Osteichthyes.
A mineralogia apontou composição predominante de apatita nas estruturas ósseass, donde se conclui que os processos de fossilização (estratinômicos e diagenéticos) impostos aos ictiólitos não alteraram a composição química original dos esqueletos, como pontuado por Martill (1988, 1989).
Finalmente, a presença de barita abre espaço para questionamentos a respeito das reais condições de oxigenação reinantes durante a fase de soterramento dos espécimes, como atribuídas por vários autores (entre eles Maisey, 1991). Assim e a propósito, permanecem as controvérsias a respeito das condições ambientais em que os peixes foram soterrados.
Agradecimentos
Os autores agradecem a todos os que de uma ou outra maneira contribuíram para a feitura do trabalho. Ao Departamento de Geologia da Universidade Federal do Pará pela cessão do material de acervo utilizado nas análises. Ao prof. Dr. Marcondes Costa pelo empréstimo da amostra 560-V, que ilustra esse trabalho. Ao Museu Paraense Emílio Goeldi, nas pessoas do Dr. Horácio Higushi, pelas importantes contribuições de caráter taxonômico e do Dr. Hilton Costi, pelas análises por EDS/MEV realizadas.
Referências
Beurlen, K. A Geologia da Chapada do Araripe. Anais da Academia Brasileira de Ciências. 34 (3): 365 – 370. 1962
________– A Paleontologia do Cretáceo no Nordeste do Brasil. Anais da Academia Brasileira de Ciências. 43 (supl.): 89 – 101. 1971
Bonfim Júnior, F. C. A Apatita na Composição dos Dentes e na Fosfatização de Tecidos Moles em Fósseis de Vertebrados – Revista Brasileira de Paleontologia 2: 74 – 75.
Brito, I. M. Nota preliminar sobre os insetos da Formação Santana, Cretáceo Inferior da Chapada do Araripe. Anais do XXXIII congresso brasileiro de Geologia. Rio de Janeiro, RJ. pp. 530 – 535. 1984
_________ Breve histórico sobre a estratigrafia da bacia do Araripe, Nordeste do Brasil. Atas do Atas do I Simpósio sobre a bacia do Araripe e bacias interiores do Nordeste. Cidade do Crato, CE. pp. 1 – 18. 1990
Brito, P. M. O Endocrânio de Tharrhias (TELEOSTEI, GONORHYNCHIFORMES) do Cretáceo Inferior da Chapada do Araripe, Nordeste do Brasil. Atas do Atas do I Simpósio sobre a bacia do Araripe e bacias interiores do Nordeste. Cidade do Crato, CE. pp. 375 – 383. 1990
Campos, D. A.; Wenz, D. Première découverte de Coleacanthes dans le Crétacé inferieur de la Chapada do Araripe (Brésil). C. R. Acad. Sc. Paris, 294 (Série II): 1151 – 1154. 1982
Carroll, R. L. Vertebrate Paleontology and evolution. W. H. freeman and Company. New York, NY. 698pp. 1988.
Costa, M.L. Mineralização de fragmentos de tecidos ósseos fósseis de sedimentos da Bacia do Acre. In: Klein, E. et al. Contribuições a Geologia da Amazônia, v.3, p. 301-318. 2002.
Lima, M. R. O Paleoambiente Deposicional da Formação Santana (Grupo Araripe) segundo evidências Palinológicas. Anais do XXX Congresso Brasileiro de Geologia. Recife, PE. 2: 970 – 974. 1978
_________ Considerações sobre a subdivisão estratigráfica da Formação Santana, Cretáceo do Nordeste do Brasil. Revista brasileira de Geociências, São Paulo, 9 (2): 116 – 121. 1979
Mabesoone, J. M.; Tinoco, I. M. Paleoecology of the Aptian Santana Formation (Northeasthern Brazil). Palaeogeography, Palaeoclimatology, Paleontolgy. 14: 87 – 118. 1973
Maisey, J. G Santana Fossils: An Illustrated Atlas. Neptune, N.J.: TFH Publications, 459 pp. . 1991
Martill, D. M. Preservation of fishes in the Cretaceous Santana Formation of Brazil. Palaeontology, 31: 1 – 9. 1988
___________The Medusa effect: instantaneous fossilization. Geology today. Nov. – Dec. 1989: 201 – 205. 1989
Martins-Neto, R. G.; Santos, J. C. K.; Mesquita, M. V.A Paleoentomofauna brasileira: Estado da Arte. XIV Simpósio de Geologia do Nordeste. Boletim 12: 59 – 62. 1991
Martins-Neto, R. G.; Castilho-Dias, C.; Fragoso, L. M. de C.; Santos, J. C. K.; Vieira, F. R. de M. II Simpósio sobre as Bacias Cretácicas Brasileiras – Resumos expandidos. Rio Claro, SP. pp. 65 – 67. 1992
Santos, M. E. C. M. Ambiente deposicional da Formação Santana – Chapada do Araripe (PE/PI/CE). Anais do XXXII Congresso Brasileiro de Geologia. Salvador, BA. 4: 1413 – 1419. 1982
_____________ Fósseis do Nordeste do Brasil: Paleoictiofáunula da Chapada do Araripe. Rio de Janeiro, RJ. Ed. da Universidade do Estado do Rio de Janeiro. 64pp. 1991
Small, H. Geologia e Suprimento d’Água Subterrânea no Ceará e parte do Piauhy. Inspeção de Obras contra Seccas, Ser. I. D. Rio de Janeiro, RJ. N. 25, 80 pp. 1913
Wenz, S.; Brito, P. M L’Ichthyofaune des nodules fossiliferès de la Chapada do Araripe (N – E du Brésil). Atas do I Simpósio sobre a bacia do Araripe e bacias interiores do Nordeste. Cidade do Crato, CE. 337 – 349. 1990